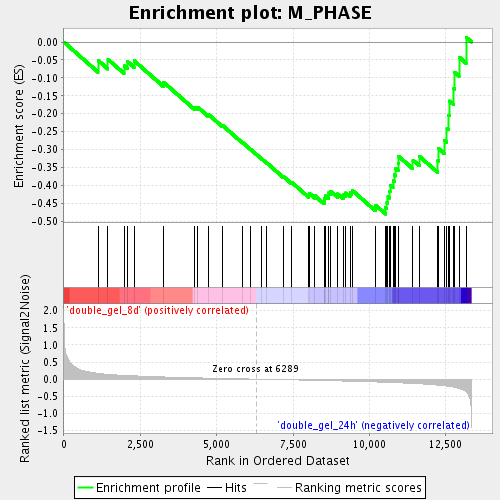
Profile of the Running ES Score & Positions of GeneSet Members on the Rank Ordered List
| Dataset | double_gel_and_single_gel_rma_expression_values_collapsed_to_symbols.class.cls #double_gel_8d_versus_double_gel_24h.class.cls #double_gel_8d_versus_double_gel_24h_repos |
| Phenotype | class.cls#double_gel_8d_versus_double_gel_24h_repos |
| Upregulated in class | double_gel_24h |
| GeneSet | M_PHASE |
| Enrichment Score (ES) | -0.48015258 |
| Normalized Enrichment Score (NES) | -1.630746 |
| Nominal p-value | 0.0073710075 |
| FDR q-value | 0.13787214 |
| FWER p-Value | 1.0 |
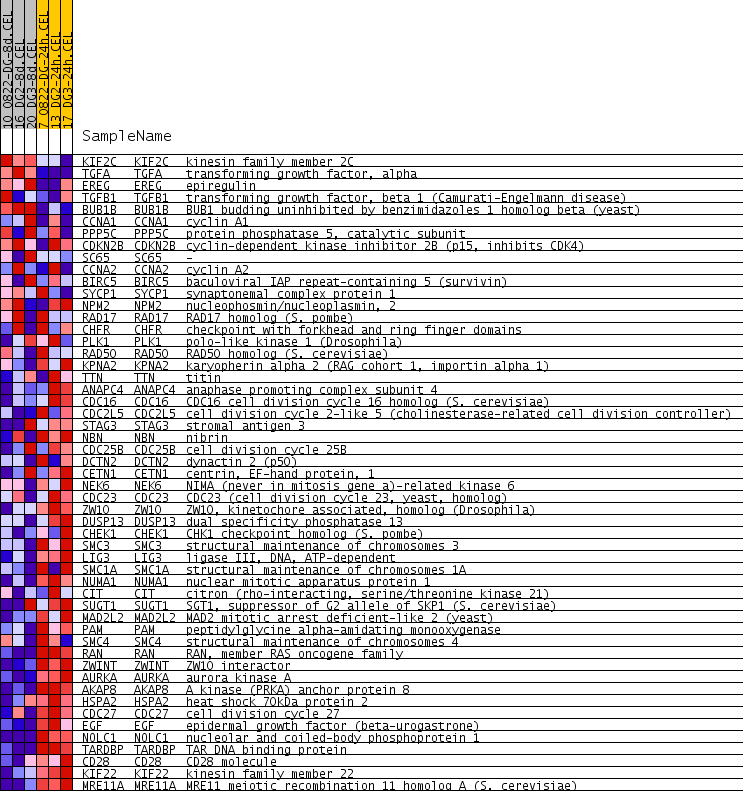
| PROBE | GENE SYMBOL | GENE_TITLE | RANK IN GENE LIST | RANK METRIC SCORE | RUNNING ES | CORE ENRICHMENT | |
|---|---|---|---|---|---|---|---|
| 1 | KIF2C | KIF2C Entrez, Source | kinesin family member 2C | 1116 | 0.169 | -0.0510 | No |
| 2 | TGFA | TGFA Entrez, Source | transforming growth factor, alpha | 1441 | 0.140 | -0.0481 | No |
| 3 | EREG | EREG Entrez, Source | epiregulin | 1968 | 0.109 | -0.0664 | No |
| 4 | TGFB1 | TGFB1 Entrez, Source | transforming growth factor, beta 1 (Camurati-Engelmann disease) | 2076 | 0.105 | -0.0540 | No |
| 5 | BUB1B | BUB1B Entrez, Source | BUB1 budding uninhibited by benzimidazoles 1 homolog beta (yeast) | 2298 | 0.096 | -0.0520 | No |
| 6 | CCNA1 | CCNA1 Entrez, Source | cyclin A1 | 3266 | 0.064 | -0.1124 | No |
| 7 | PPP5C | PPP5C Entrez, Source | protein phosphatase 5, catalytic subunit | 4288 | 0.038 | -0.1818 | No |
| 8 | CDKN2B | CDKN2B Entrez, Source | cyclin-dependent kinase inhibitor 2B (p15, inhibits CDK4) | 4378 | 0.036 | -0.1814 | No |
| 9 | SC65 | SC65 Entrez, Source | - | 4719 | 0.030 | -0.2012 | No |
| 10 | CCNA2 | CCNA2 Entrez, Source | cyclin A2 | 5186 | 0.020 | -0.2322 | No |
| 11 | BIRC5 | BIRC5 Entrez, Source | baculoviral IAP repeat-containing 5 (survivin) | 5848 | 0.008 | -0.2804 | No |
| 12 | SYCP1 | SYCP1 Entrez, Source | synaptonemal complex protein 1 | 6106 | 0.003 | -0.2991 | No |
| 13 | NPM2 | NPM2 Entrez, Source | nucleophosmin/nucleoplasmin, 2 | 6472 | -0.003 | -0.3260 | No |
| 14 | RAD17 | RAD17 Entrez, Source | RAD17 homolog (S. pombe) | 6622 | -0.006 | -0.3360 | No |
| 15 | CHFR | CHFR Entrez, Source | checkpoint with forkhead and ring finger domains | 7176 | -0.015 | -0.3746 | No |
| 16 | PLK1 | PLK1 Entrez, Source | polo-like kinase 1 (Drosophila) | 7458 | -0.021 | -0.3917 | No |
| 17 | RAD50 | RAD50 Entrez, Source | RAD50 homolog (S. cerevisiae) | 8016 | -0.032 | -0.4275 | No |
| 18 | KPNA2 | KPNA2 Entrez, Source | karyopherin alpha 2 (RAG cohort 1, importin alpha 1) | 8037 | -0.032 | -0.4228 | No |
| 19 | TTN | TTN Entrez, Source | titin | 8210 | -0.035 | -0.4288 | No |
| 20 | ANAPC4 | ANAPC4 Entrez, Source | anaphase promoting complex subunit 4 | 8515 | -0.042 | -0.4435 | No |
| 21 | CDC16 | CDC16 Entrez, Source | CDC16 cell division cycle 16 homolog (S. cerevisiae) | 8534 | -0.042 | -0.4366 | No |
| 22 | CDC2L5 | CDC2L5 Entrez, Source | cell division cycle 2-like 5 (cholinesterase-related cell division controller) | 8548 | -0.042 | -0.4293 | No |
| 23 | STAG3 | STAG3 Entrez, Source | stromal antigen 3 | 8660 | -0.045 | -0.4290 | No |
| 24 | NBN | NBN Entrez, Source | nibrin | 8672 | -0.045 | -0.4210 | No |
| 25 | CDC25B | CDC25B Entrez, Source | cell division cycle 25B | 8722 | -0.046 | -0.4158 | No |
| 26 | DCTN2 | DCTN2 Entrez, Source | dynactin 2 (p50) | 8962 | -0.051 | -0.4238 | No |
| 27 | CETN1 | CETN1 Entrez, Source | centrin, EF-hand protein, 1 | 9141 | -0.055 | -0.4265 | No |
| 28 | NEK6 | NEK6 Entrez, Source | NIMA (never in mitosis gene a)-related kinase 6 | 9217 | -0.056 | -0.4212 | No |
| 29 | CDC23 | CDC23 Entrez, Source | CDC23 (cell division cycle 23, yeast, homolog) | 9365 | -0.060 | -0.4206 | No |
| 30 | ZW10 | ZW10 Entrez, Source | ZW10, kinetochore associated, homolog (Drosophila) | 9428 | -0.061 | -0.4133 | No |
| 31 | DUSP13 | DUSP13 Entrez, Source | dual specificity phosphatase 13 | 10198 | -0.082 | -0.4552 | No |
| 32 | CHEK1 | CHEK1 Entrez, Source | CHK1 checkpoint homolog (S. pombe) | 10530 | -0.091 | -0.4624 | Yes |
| 33 | SMC3 | SMC3 Entrez, Source | structural maintenance of chromosomes 3 | 10571 | -0.092 | -0.4474 | Yes |
| 34 | LIG3 | LIG3 Entrez, Source | ligase III, DNA, ATP-dependent | 10605 | -0.094 | -0.4316 | Yes |
| 35 | SMC1A | SMC1A Entrez, Source | structural maintenance of chromosomes 1A | 10658 | -0.096 | -0.4168 | Yes |
| 36 | NUMA1 | NUMA1 Entrez, Source | nuclear mitotic apparatus protein 1 | 10675 | -0.096 | -0.3993 | Yes |
| 37 | CIT | CIT Entrez, Source | citron (rho-interacting, serine/threonine kinase 21) | 10772 | -0.100 | -0.3870 | Yes |
| 38 | SUGT1 | SUGT1 Entrez, Source | SGT1, suppressor of G2 allele of SKP1 (S. cerevisiae) | 10813 | -0.101 | -0.3702 | Yes |
| 39 | MAD2L2 | MAD2L2 Entrez, Source | MAD2 mitotic arrest deficient-like 2 (yeast) | 10862 | -0.103 | -0.3537 | Yes |
| 40 | PAM | PAM Entrez, Source | peptidylglycine alpha-amidating monooxygenase | 10944 | -0.106 | -0.3390 | Yes |
| 41 | SMC4 | SMC4 Entrez, Source | structural maintenance of chromosomes 4 | 10956 | -0.107 | -0.3190 | Yes |
| 42 | RAN | RAN Entrez, Source | RAN, member RAS oncogene family | 11423 | -0.126 | -0.3295 | Yes |
| 43 | ZWINT | ZWINT Entrez, Source | ZW10 interactor | 11637 | -0.135 | -0.3191 | Yes |
| 44 | AURKA | AURKA Entrez, Source | aurora kinase A | 12238 | -0.175 | -0.3302 | Yes |
| 45 | AKAP8 | AKAP8 Entrez, Source | A kinase (PRKA) anchor protein 8 | 12267 | -0.177 | -0.2978 | Yes |
| 46 | HSPA2 | HSPA2 Entrez, Source | heat shock 70kDa protein 2 | 12445 | -0.193 | -0.2735 | Yes |
| 47 | CDC27 | CDC27 Entrez, Source | cell division cycle 27 | 12536 | -0.203 | -0.2407 | Yes |
| 48 | EGF | EGF Entrez, Source | epidermal growth factor (beta-urogastrone) | 12591 | -0.209 | -0.2039 | Yes |
| 49 | NOLC1 | NOLC1 Entrez, Source | nucleolar and coiled-body phosphoprotein 1 | 12621 | -0.214 | -0.1644 | Yes |
| 50 | TARDBP | TARDBP Entrez, Source | TAR DNA binding protein | 12761 | -0.235 | -0.1291 | Yes |
| 51 | CD28 | CD28 Entrez, Source | CD28 molecule | 12792 | -0.240 | -0.0846 | Yes |
| 52 | KIF22 | KIF22 Entrez, Source | kinesin family member 22 | 12933 | -0.273 | -0.0418 | Yes |
| 53 | MRE11A | MRE11A Entrez, Source | MRE11 meiotic recombination 11 homolog A (S. cerevisiae) | 13163 | -0.372 | 0.0135 | Yes |